
Aquaporins (AQPs): On Making Progress Towards their Role and Application in the Biological World
-
Pratiksha Upadhyay
Faculty of Science, Haflong Government College, Assam University, Haflong-788819, India
Rishikesh Upadhyay

Faculty of Science, Haflong Government College, Assam University, Haflong-788819, India
| Received 13 Jul, 2025 |
Accepted 29 Sep, 2025 |
Published 30 Sep, 2025 |
Aquaporins are small pores or channels about which only limited information has been documented. Despite their small size, they are believed to play significant roles in the body, particularly within the plasma or cell membranes of both plant and animal cells. These channels have been identified not only in plants but also in various animal tissues and cells. However, comprehensive data on them remains scarce. Through this review article, it is intended that increased awareness be generated among researchers and the scientific community regarding the presence of aquaporins in the biological world, their essential functions in plant and animal systems, and their potential significance in future research. In this paper, relevant information has been compiled concerning the history, development, structure, functions, and distribution of aquaporins. Furthermore, their physiological and pathological roles in both plant and animal cells have been explored and discussed.
INTRODUCTION
The word aquaporins begins from the two Latin words “aqua”, meaning “water”, and “porus”, meaning little opening, pores, or channels. The aquaporins are the individuals from those essential layer protein super families that structure little channels to work with the quicker transportation or trade of water and other little solutes in the middle of between the cells or natural films of practically all living organic entities, including plants and creatures, across their endothelial and epithelial boundaries. They are those family members of water proteins which are responsible for regulating the osmotic pressure in the body. But it is to be noted that the water transporting capacity of aquaporin monomer is very low so membranes are designed to contain lots of aquaporins monomers in order to have faster and more water transportation facility. Aquaporins in plants are recognized for their fundamental roles in maintaining turgor pressure, regulating root and leaf hydraulics, facilitating leaf transpiration, enabling water uptake in roots, and mediating responses to various biotic and abiotic stresses1,2. In addition to water transport, these membrane proteins are also involved in the movement and exchange of volatile substances such as CO2, NH3, and metalloids, including boron, silicon, and germanium. Plant aquaporins have been localized in various cellular compartments such as the plasma membrane, tonoplast, endoplasmic reticulum, vacuoles, and plastids. Similarly, in animals, aquaporins are considered integral components of physiological systems. Their primary function is to maintain homeostasis through the regulation of water, ion, and solute transport. In certain contexts, they are also responsible for the transfer or exchange of metals and other chemicals across cellular membranes3-5. Key physiological roles are fulfilled by aquaporins in animals, including the regulation of cell volume, facilitation of diffusion, and participation in essential processes such as kidney function (urine concentration), saliva secretion, moisture retention in the respiratory system, and water balance in the brain. Additionally, their involvement has been observed in apoptosis, cell migration, and mitochondrial metabolism. These proteins have been identified in various tissues, such as the lens (to maintain hydration and transparency), proximal tubules, and descending loops (to facilitate water reabsorption). At the molecular and cellular levels, the regulation of aquaporins in both plant and animal cells has been shown to respond to signaling intermediates, including cytosolic pH, calcium ions, and Reactive Oxygen Species (ROS). The objective of this research is to explore the comparative regulatory mechanisms and physiological significance of aquaporins across plant and animal systems. By doing so, the study aims to highlight conserved and divergent functions, contributing to a more integrated understanding of membrane transport biology. The significance of this investigation lies in its potential to bridge plant and animal physiology, expanding applications in biotechnology, agriculture, and medicine. Ultimately, new knowledge is advanced by identifying novel regulatory pathways and multifunctional roles of aquaporins, thereby enhancing our understanding of water and solute dynamics in living systems.
HISTORY AND EVOLUTION
The tribute goes to Dr. Peter Agre, an American biochemist, and his research team, who identified and characterized the first Aquaporin protein in red blood cells of humans in 1996. They named this protein CHIP28 (channel-forming integral protein of 28 kilodaltons), which subsequently came to be known as Aquaporin-1(AQP1). The Aquaporins belong to a highly conserved superfamily arche type of archetypal membrane proteins called Major Intrinsic Protein (MIP) known to be present in the mammalian lens fibers, which is very huge as well as complex in its own constitution and includes more than 150 membrane channel proteins it. This ability of Aquaporins shows that they facilitate transport of complex substances like urea, glycerol, ammonia, ions, CO2, O2, boron, antimony, arsenic, silicon, H2O2, etc., and monitor the water transportation system among the plant and animal cells to help them adapt themselves in the harsh environment, apart from assisting them in carrying out their vital activities. However, in the case of animal cells, these aquaporins are tissue or organ-specific and can be regulated by hormones. It is worth mentioning here that the amount of aquaporins as well as their isoforms, in plant cells is much greater than in animal cells.
The family of Aquaporin is diverse in its own and its members are present throughout all the kingdoms of life, such as Archaea, Eubacteria, Fungi, Plants, and Animals. As mentioned earlier, there is diversity in the family. So, those MIPs that are extremely permeable to water exchange or transport are the ones known as Aquaporins. Likewise, they are permeable to glycerol and water, called Aquaglyceroporins. The APQs are very sensitive to mercury, and this is the earliest evidence against the protein involvement in transmembrane water access3.
Plant aquaporins: The plants’ MIPs arise as multiple isoforms in nature and are classified into seven subfamilies based on their subcellular localizations and other specific features. Among them, five are found in seed plants: Plasma membrane intrinsic proteins (PIPs), tonoplast intrinsic proteins (TIPs), NOD26-like intrinsic proteins (NIPs), small basic intrinsic proteins (SIPs), and unknown intrinsic proteins (XIPs). The MIPs, like hybrid intrinsic proteins (HIPs) and GLpF- like intrinsic proteins (GIPs) are present only in older mosses. GIPs are the only GLPs to be present in plants. As discussed earlier, the different plant groups contain different numbers of Aquaporin isoforms (AQI). For instance, the rice place contains 33 AQI while maize, tomato, and soybean has 31, 47 and 66 of them respectively4. Ample reasons lie behind this due to the higher duplication rates of plants in their genomes, their static lifestyle, and the adaptability
of the aquaporins, and most importantly, the aquaporins in the case of plants are very species specific. The water shipping ability of the plant Aquaporins was witnessed for the first time in Arabidopsis sp. One Aquaporin, PM28A, and two Aquaporin homologous, PM28B, and so-dTIP were identified in spinach. PM28A and PM28B are located in the plasma membrane, where they constitute approximately 15% of the total protein, whereas the so-dTIP is located in the vacuolar membrane. Studies confirm that there are specific aquaporins present in plants for the pollens. For instance, TIP1; 3, TIP5; 1, NIP4; 1 and NIP4; 2 are some of the pollen-specific aquaporins present in plants. Unlike other AQPs, these exhibit low water permeability and higher transportation of substances like glycerol, urea, and are even found involved in the nitrogen utilization pathway during the growth period of pollen tube5. After the discovery of plant aquaporin, the earlier view that no protein-mediated water infiltration is needed in the plants was highly challenged. After that, it is revealed that steroid hormones are involved in the homeostasis regulation of aquaporins. The aquaporins take part only in the transcellular pathway in case of the organs of plant and not in the apoplastic and symplastic pathways for water transportation. It is seen that the transcellular transportation occurs at first in endodermis due to the presence of the plasma membrane aquaporins. In the case of a plant, it is seen that some APQs remain there in the plant body till the growth process after that they used to disappear, while some other continue their expression6.
Animal aquaporins: After extensive research, Scientists discovered that the 3D structure of the human Aquaporin AQP1 is almost similar to the bacterial Aquaglyceroporin GlpF with respect to its structure, except the 30% difference in the two sequence arrangements at the amino- acid- level between them. This shows that the human Aquaporins have evolved from the bacterial glyceroporins at some points. Animal MIPs are generally classified into four broad groups: classical or orthodox AQPs (AQP0, 1, 2, 4, 5, 6, 14) associated with water transport, aqua-ammonioporins (AQP8), the Aquaglyceroporins (AQP3, 7, 9, 10, 13), and the unorthodox AQPs (also superaquaporins) (AQP11-12)7.
In mammals, there are 13 distinct types of aquaporins identified, each with 12-15 isoforms (AQP0-12), corresponding to human aquaporin proteins ranging from AQP0 to AQP10. However, human aquaporins exhibit 18 paralogs due to tandem duplications, including four additional AQP7 pseudo genes and a duplicated AQP12 gene. Among these, aquaporins such as AQP3, AQP7, AQP9, and AQP10 are known to be involved in glycerol transport and are classified under the GLP (Glycerol Permeability) subfamily. The aquaporin genes found in humans have four to eight introns it, and the size of the gene varies between 3.6 kilobases (kb) to 47 kb. They have been successfully mapped to some specific chromosomes, including 1, 7, 9, 10, 12, 15, and 16 chromosomes. Notably, the genes encoding AQP0, AQP2, AQP5, and AQP6 are clustered on chromosome 12. Additionally, splice variants have been identified for the genes encoding human AQP1, AQP4, and AQP68.
It is seen that the aquaporins in human exhibit their different roles through cells and tissues of the body. For e.g., the AQP0 is found mostly in the lens, AQP1 itself in many organs like kidneys, liver, pancreas, vascular endothelial cells, skin, heart, and brain9, AQP2 in the collecting tubules of the kidneys. Although AQP1, 2, 3, and 4 are all found to be present in the kidneys but the controlling mechanism of AQP2 is familiar to us till now. In research conducted on the rat model, it has been found that an excess of APQ2 expression can cause congestive heart failure. The AQP3 is present in the skin10 and in the case of sperm, both AQP3 and 7 are present. The AQP4 is also found in the brain, specifically in the plasma membrane of astrocytes, the most abundant glial cells of the brain, and in the neuromyelitis optica. Similarly, AQP9 is found to be present in the epididymis11.
Recent studies show that any type of mutation in the AQP2 can lead to metabolic disorders like diabetes insipidus in humans. Some studies reveal the involvement of various AQPs in the metastatic tumors’ growth and development, leading to the formation of cancer also and their deletion leads to the opposite can be seen in the implanted tumors during one experiment12.
Structure: In nature, the Aquaporins occur usually as tetramers and all of their monomers depict a specific type of arrangement of six transmembrane-straddling alpha helices (1-6), joined by three extracellular (i.e., loop A, C, and E) and two intracellular loops (i.e., loop B and D)13. On top of that, both the N and C termini are located on the cytoplasmic side. In case of animal cells, especially mammalian ones, the homotetrameric association of aquaporins whereas in case of plant cells, the heterotetrameric association can be studied generally. Each of its monomers operates a separate water channel, exhibiting an independent nature among them. Both B and E loop forms two short hydrophobic alpha helices separately, sinking halfway into the membrane from the opposite sides of the Aquaporin. Thus, the forming pores of high specificity emerge from two filter regions. The first filter region is formed when the highly conserved amino acid, namely Asparagine-Proline-Alanine (NPA) motif of loop B and E, meets at the core region of the channel and forms a primary size exclusion zone. Similarly, the second one, also called Arg, is formed when the four amino acids located in the extracellular region of the pore come together to form the size exclusion barrier and hydrogen bond for the transport of substrate molecules13.
EFFECT OF DIFFERENT TYPES OF STRESS ON PLANT AND ANIMAL AQUAPORINS
Water stress: Based on the evidence gathered till now, it is widely accepted that the process of intake of water as well as the intercellular water flow in roots, is mediated by the two most abundant aquaporins present in the plasma membrane, namely, PIPs and TIPs. After many studies, the scientist came to the conclusion that during the time of drought stress, except AtPIP1; 4 and AtPIP2; 5 all other PIP aquaporins tend to get down-regulated in the leaves of plants. In various parts of different plants, such as the fruit of the peach plant, twigs of olive plants, etc, sudden but powerful downregulation of PIP genes can be seen during some adverse conditions, like drought. Here it is to be mentioned that in case of different cultivars of the same plant species, the transcriptional expressions as well as the responses of the same homologous aquaporin family differ distinctly toward drought stress. For instance, unlike an isohydric cultivar, in the case of anisohydric one, upregulation of VvPIP1; 1 gene can be observed in the roots of grapevines during drought stress conditions.
For a second time in another experiment, shockingly, down-regulation of VvPIP2; 1 aquaporins can be seen during drought conditions. Therefore, very contrasting results have been observed between similar aquaporin homologs within the same species under drought stress. Reported that arbuscular mycorrhizal symbiosis with maize plants can control the various expressions of aquaporins differently during both temporary and extended drought stress conditions14. Studies conducted in naturally drought-tolerant plant species to approximate aquaporins role during drought conditions. A drought-tolerant plant, Eragrostis nindensis (resurrection grass), has TIP3; 1 in small vacuoles of its leaves, which plays a major role by improving the mobilization of water and solute molecules from those small vacuoles after rehydration. The transgenic plant shows better resistance and enhanced growth and development when subjected to drought stress. Animal AQPs also played a huge role in the homeostasis of water balance in the animal body. The AQPs found in the kidney, mainly the AQP2, are associated with osmoregulation and water homeostasis. They do so by activating the ADH hormone during water-deficient conditions, reabsorbing large amounts of water, and producing very concentrated urine as output15.
Salt stress: However much salinity there is in the soil, it is due to the combined effects of the water and the ionic salts, which cause toxic effects on plant growth and development by disturbing their cells’ osmotic balance. The primary responses of plants towards the salt stress are the inhibition of root-water uptake and resultant decrease in the levels of Lpr, also observed in the case of the drought stress response. So, from a variety of experimental evidence, it can be summarized that the response of aquaporins in the context of salinity varies with drought stress. For example, in Arabidopsis, a significant reduction in Lpr, along with 60% to 75% decrease in PIP and TIP aquaporin transcripts abundance, was observed after its exposure to salt stress. Several transgenic banana plants overexpressing MusaPIP1; 2 and MusaPIP1; 2exhibits enhanced tolerance to both salt and drought stress. The transcript and protein levels in the case of the barley HvPIP2; 1 gene were found to be down-regulated in roots but up-regulated in the shoot portion of plants under salt stress. Several studies involving the localization of aquaporins during salt stress conditions in plants have revealed re-localization or redistribution of aquaporins in response to high salt concentrations16. For example, the salt stress-induced re-localization of AtTIP1; 1 into intravacuolar invaginations is seen in Arabidopsis. Apart from this, an alternative mechanism regulating the PIP abundance in the plasma membrane i.e., endocytosis of PIPs either through the clathrin-dependent pathway or a salt- stimulated membrane raft–associated pathway, can also be observed in Arabidopsis. Moreover, the relocalization of TIP1; 1-GFP into intracellular spherical structures and internalization of PIP2; 1-GFP in response to salt stress were observed in Arabidopsis. The occurrence of both endocytosis and exocytosis, triggered by the cycling of AtPIP2; 1 to and from the plasma membrane, is also noted during exposure to salt stress. Animal aquaporins (AQPs) are essential during salt stress as they facilitate the movement of water across cell membranes, helping to maintain cellular water balance in challenging conditions. They help regulate water flow, which is vital for mitigating the impact of salt stress on cellular function and overall organismal physiology2.
Cold stress: Like other abiotic stresses, it also plays a crucial role in the growth of plants during their development1. It’s been found that the overexpression of the cold stress controlling aquaporins i.e., MusaPIP1; 2 demonstrates tolerance towards both cold and drought stress in transgenic banana plants. Unlike non-transgenic tobacco plants, the transgenic ones contain a wheat aquaporin TaAQP7 (PIP2) gene which, during its overexpression, provides tolerance to the plant against both cold and drought stresses. What is surprising is the above case does not match with all the plants. The contrasting results were noticed in various experiments on variety of plant species. For example, the transgenic Arabidopsis plants that contain aquaporins like AtPIP1; 4 and AtPIP2; 5 during their overexpression, exhibited tolerance toward cold stress but became very prone to the drought stress due to frequent water loss. Research finding reveals that in the case of sensitive plants like rice, the level of Lpr increases when the plants are exposed to cold stress for a long time. It is also found that some aquaporins like OsTIP1; 1 and OsTIP2;2 get drastically down-regulated during cold stress, while aquaporins like OsPIP1;3 tend to get up regulated by 60% in roots region due to the cold stress17. During the cold stress in animals, aquaporins (AQPs) play a crucial role in regulating water movement and maintaining cellular balance. Specifically, AQP3 and AQP9 are recognized for their involvement in responses to cold stress. The AQP3 facilitates glycerol transport, which is essential for preserving cell membrane fluidity, apart from preventing cellular damage caused by ice formation in cold conditions. Recent research, such as that conducted by Li et al., explored the role of AQP3 in the liver of the plateau pika (Ochotona curzoniae), revealing that AQP3 expression increases in response to cold exposure. This upregulation facilitates glycerol transport, contributing to the animal’s ability to tolerate cold environments. The AQP9 is another aquaglyceroporin known for its role in glycerol and small solute transport across membranes. It also contributes to glycerol metabolism and cold adaptation in various species, including fish and mammals.
Heavy metal stress: In nature, many metals exist and are proved to be beneficial or crucial subject to their limited existence for the development of plants. It is well known that the aquaporins not only facilitate the transport of water but also act as a channel membrane for the transportation of some heavy metals like B, Si, Sb, As., etc. These vary metals are the ones which, if remained or given in a specific amount to plants, can successfully stimulate their growth and development. However, any excess can give stress to both plants and animals, called heavy metal stress18,19. In plants, the metals like B and Si seem to be very useful for their growth and development. At the same time, metals like As, Sb etc prove to be very toxic for them. Some knockout mutant aquaporins like AtNIP6; 1, AtNIP7; 1 exhibit an involvement in the transportation and tolerance against heavy metals like arsenic (As) and antimony (Sb). Their up and down regulations help in detoxifying these vary elements in plants. The high level of tolerance against arsenic (As) stress is shown by the double knock out mutants of plant aquaporins like AtNIP1; 1 and AtNIP3; 1. The mutant of AtNIP1; 1 also shows tolerance against extreme Sb stress. Animal aquaporins (AQPs) also play vital roles in managing cellular homeostasis during heavy metal stress. Specifically, AQP7 and 9 facilitate the movement of small solutes, including heavy metals like arsenite, aiding detoxification by exporting or sequestrating these metals. Additionally, AQP8 helps mitigate oxidative stress by transporting hydrogen peroxide (H2O2), protecting cells from oxidative damage. Furthermore, these AQPs are involved in signaling pathways that activate cellular defense mechanisms in response to heavy metal exposure20.
ROLE OF PLANT AND ANIMAL AQUAPORINS IN PHYSIOLOGY AND PATHOLOGY
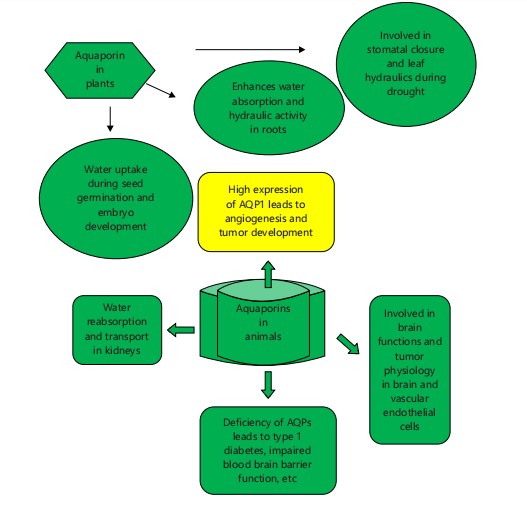
Aquaporins are integral to the physiological and pathological processes in both plant and animals (Fig. 1). In plants, AQPs are crucial for water uptake during seed germination and embryo development, as evidenced in species such as Arabidopsis thaliana and Vicia faba21. In roots, AQPs enhance water absorption and hydraulic conductivity, with studies indicating reductions up to 47% and 64% in Populus and Arabidopsis, respectively, when AQP function is inhibited22. Additionally, AQPs are involved in stomatal closure and leaf hydraulics, responding to environmental stresses like drought23. In animals, AQPs are found in various organs, including the kidney, brain, and pancreas. For example, AQP1 is prevalent in the kidney and plays a role in water reabsorption and the transport24. The AQPs are also present in the brain and vascular endothelial cells, contributing to brain functions and tumor physiology. High AQP1 expression in tumors is linked to tumor development and angiogenesis25. Deficiencies in AQPs can lead to disorders such as diabetes insipidus and impaired blood-brain barrier function, emphasizing their pathological importance26. Notably, AQP4 is associated with neuromyelitis optica, an autoimmune disease affecting water transport in the brain27.The animal aquaporins, particularly AQP3, 7, 9 and 10 are crucial in the pathological processes related to the heavy metal stress and transport. These aquaporins facilitate the diffusion of trivalent arsenite (similar to glycerol) and antimonite, both toxic compounds. Arsenite enters cells via these aquaporins and is involved in chemotherapy for treating certain leukemia types and protozoan parasitic diseases. Antimonite, transported similarly, is toxic and can enter the food chain. used to treat diseases like leishmaniasis and some parasitic infections and types of leukemia. Increased expression of specific aquaporins in human leukemia cells leads to hypersensitivity to antimonite due to higher accumulation levels28.
ROLE OF AQUAPORINS IN VARIOUS TYPES OF HUMAN CANCER
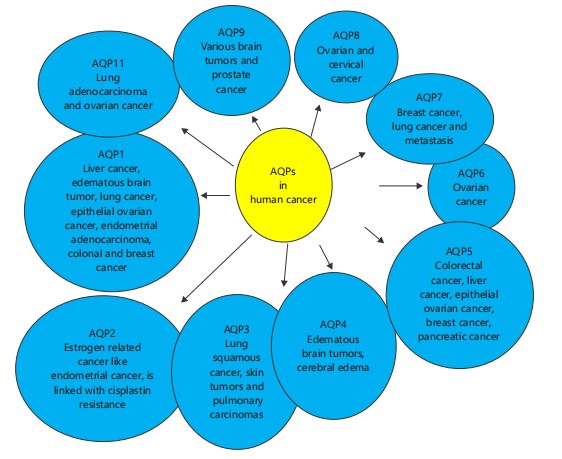
It is known that the aquaporins are those tiny channel proteins that facilitate the water transport along with some solutes in both plants and animals. But it is to be mentioned that just like everything has some kind of dark side, they also have. The overexpression of all types of aquaporins can lead to carcinogenic effects in the bodies of both plants and animals. They do so by actively taking part in metastasis, cell growth, and proliferation, etc. Each type of aquaporin exerts different methods or way to show their effect in different kinds of cancer29. For e.g., some aquaporins like APQ11 and 12 are those super aquaporins which are found inside the cytoplasm to facilitates water transport and AQP11, mainly found in ER, its expression is seen to be involved in the ovarian cancer patients. After a research conducted it is found that the signaling process like EGF and EGFR-mediated signaling pathways or pathways similar to them are involved in the process like metastasis, migration of tumor cell etc. Furthermore, role of various AQP expression in the lung, brain tumors, have been predicted. In the late stages of metastatic liver the expression of AQP1 and 5 has been found. Similarly from the early to the late stages of the colorectal cancer the expression of AQP5 can be seen. Various studies propose that the AQP1 and 4 find a key role in the edematous brain tumors. In case of metastatic cancers, the expression of AQP1 is found to be present in areas such as microvessel endothelia30. The AQP1 also participates in the creation and development of brain tumor edema. In the case of epithelial ovarian tumors, the relationship between the expression of AQP5 and 1 can be seen. After the studies or researches conducted by Pan et al.31, the expression of AQP1 in the case of the growth and development of endometrial adenocarcinoma can be established.
|
In between the metastatic carcinomas, the expression of AQP is seen in glioblastoma cells and not in the parenchyma or normal micro vessel endothelium of the normal brain32. Jiang verified that the expression of AQP1 can be found in quite a few cancer cell lines and also in a colon cancer model of human HT2033. It is found that in case of breast cancer an increased expression of AQP1, 3 and 5 has been found. It is seen that during the advancement of particular lung squamous cell cancer the expression of AQP3 increases and not during the adeno carcinoma cells. As the AQP3 are predominantly found or expressed in the human skin so in a research conducted on the mice model it is found that the mice model where the AQP3 were absent they were devoid of skin tumors and are resistant to it even after the exposure to tumor causing chemical such as phorbol ester as compared to the one in which the AQP3 were present. Alike AQP1 the AQP3 also participates in the development of pulmonary carcinomas. As AQP4 is a water channel specifically present in the brain so the involvement of its expression can be seen in the development and progression of brain-related carcinomas like brain tumors, cerebral edema etc.34. Also it controls the water flow movement in the brain in the bidirectional way so the chances of the occurrence of edema in the presence of its expression are quite common. Over expression of AQP5 can cause cancers like pancreatic cancer, colorectal cancer etc. The study report given by Woo et al., states that the overexpression of AQP5 can instigate the Ras pathway which can eventually lead to the activation of oncogenic pathway. In one study it is found that the over expression of AQP8 lead to the cervical cancer and ovarian cancer. The vigorous expression of AQP9 can be seen in case of various brain tumors. Also it has a clearance effect for glycerol and lactate, which eventually lead to the elevation of lactic acidosis in the body causing cancer.
|
The AQP9 can be exerting its expression in case of prostate cancer through the activation of signaling pathways like MMP9, ERK1. In the research report given by or received from Zou et al.35 suggests that the AQP2 expression is somewhat related to estrogen so it’s over expression can lead to estrogen related diseases or cancers like endometrial cancer. Evans et al. recommended that overexpression of AQP11 can lead to lung adenocarcinoma in humans with less survival. Out of 11 cancer cell line one study report suggest that the over expression of AQP2 is linked with cisplastin resistance36. It is found that the overexpression of AQP7 is related or the case of breast cancer as well as lung cancer or metastasis sometimes. Although there is very less information about the relation of AQP6 with any cancer but still one study reports that the overexpression of AQP6 can cause ovarian cancer (Fig. 2).
AQUAPORINS IN INFECTIOUS AND NON-INFECTIOUS DISEASES
The AQPs have their role in various infectious diseases caused by bacteria, virus, and parasites as well as non-infectious diseases like renal disease, cataract, obesity, skin diseases etc. The up-regulation of various AQPs can be seen during the infectious diseases caused by bacteria, viruses and parasites. Out of the other bacterial diseases, sepsis is the one during which the upregulation of AQP1,2, 3,5 and 9 can be seen in leucocytes, lung epithelial cells, etc in case of human, mice and rat model. In case of acute kidney injury (AKI) and endotoxin-induced AKI due to the exposure of lipopolysaccharide (LPS) in case of mouse model and rat model, the involvement of AQP1 and 2 can be seen. On the other hand AQP3 is associated with diarrhea caused by the Vibrio cholera, E. coli and C. rodentium bacteria, as well as gastric cancer caused by Helicobacter pylori bacteria37. The AQP5 is related to bronchitis and in the case of pulmonary injury caused by LPS, the upregulation of the AQP7 and 9 can be seen. Also, AQP7 is related to the diarrhea caused by E.coli and LPS along with the colonic injury caused by LPS. The AQP8 is associated with diarrhea caused by Vibrio cholera and colonic injury caused by LPS. Whereas the bacterial infection caused by Pseudomonas aeruginosa and endocarditis caused by bacterial LPS have a relationship with the upregulation of AQP9. In case of viral infections several aquaporins are associated. Some of them are: the AQP1 is associated with the liver cirrhosis caused by the hepatitis B virus, along with infantile gastroenteritis caused by rotavirus38. It is to be mentioned that in case of infantile gastroenteritis caused by rotavirus and dengue virus the upregulation of AQP4 can also be seen. The AQP 2 and 3 are associated with the cervical cancer caused by Human Papillomavirus (HPV). The AQP3 is also associated with nasopharyngeal carcinoma and lymphoma caused by Epstein-Barr virus. It is seen that HIV-associated dementia and cortical degeneration, brain edema caused by herpes simplex virus and LPS, respectively, there is with an upregulation of AQP4. The AQP8 is associated with the male subfertility caused by HPV, while AQP9 contributes to the encephalitis caused by the herpes simplex virus. Among parasitic infections, the malaria disease caused by the Plasmodium spp. shows the involvement of AQP3 and 10, while the malaria and cerebral malaria caused by Plasmodium berghei show the contribution of AQP439.
CONCLUSION
The aquaporins are a fundamental piece of the plants and creatures. Since these small diverts are available in the cells, they can do their fundamental exercises, and thus both the plants and the creatures can succeed in their everyday life. The aquaporins not only maintain the turgor pressure in the plant’s cell but also regulate homeostasis in the animal’s body. It can be said that they carry out diverse functions in the bodies of both plants and animals. The presence in plants and animals helps them battle off various kinds of stress present in their environments. There are various or diverse groups or kinds of aquaporins present in both plants and animal cells for carrying out diverse functions in different organs and tissues. For example, AQP1 and 2 are present in proximal tubules, descending loop of Henle, and collecting ducts of the kidneys, respectively, for water reabsorption. In contrast, the TIP1; 3, TIP5; 1, NIP4; 1, and NIP4; 2 are some of the pollen-specific aquaporins present in plants. From this review article, it can be known how aquaporins, like tiny water channels, are present inside the cells, which is magnificent on its own. Since it is known that the cell itself is very tiny in size, and within it, these tiny channels or pores are present. It’s very hard to believe, but it’s a fact. Many more facts about aquaporins are there that are still left to be discovered by scientists. One expectation is that such investigations will rouse a lot more researchers to carry out bunches of exploration regarding the matter so that all until now covered-up realities get uncovered in the future.
SIGNIFICANCE STATEMENT
Aquaporins (AQPs) are integral membrane proteins that facilitate the selective transport of water and small solutes across cellular membranes, playing a vital role in maintaining water homeostasis in all forms of life. Recent advances in structural biology, molecular dynamics, and functional characterization have deepened the understanding of their diverse physiological roles and regulation. This progress is paving the way for innovative applications in biotechnology, agriculture, and medicine-ranging from improving crop water-use efficiency to developing AQP-targeted therapies for edema, cancer, and neurological disorders. Hence, understanding AQPs more fully holds promise for significant breakthroughs in both basic science and applied biological systems.
REFERENCES
- Upadhyay, R., 2022. The Life of Plants in a Changing Environment. Cambridge Scholars Publishing, Newcastle upon Tyne, England, ISBN: 9781527587182, Pages: 257.
- Upadhyay, R., 2023. Stress in Plants: The Hidden Half. Cambridge Scholars Publishing, Newcastle, England, ISBN: 9781527556034, Pages: 274.
- Wayne, R. and M. Tazawa, 1990. Nature of the water channels in the internodal cells of Nitellopsis. J. Membr. Biol., 116: 31-39.
- Maurel, C., Y. Boursiac, D.T. Luu, V. Santoni, Z. Shahzad and L. Verdoucq, 2015. Aquaporins in plants. Physiol. Rev., 95: 1321-1358.
- di Giorgio, J.A.P., G.C. Soto, J.P. Muschietti and G. Amodeo, 2016. Pollen aquaporins: The solute factor. Front. Plant Sci., 7.
- Agre, P., M. Bonhivers and M.J. Borgnia, 1998. The aquaporins, blueprints for cellular plumbing systems. J. Biol. Chem., 273: 14659-14662.
- Finn, R.N. and J. Cerdà, 2015. Evolution and functional diversity of aquaporins. Biol. Bull., 229: 6-23.
- Kruse, E., N. Uehlein and R. Kaldenhoff, 2006. The aquaporins. Genome Biol., 7.
- Shields, S.D., J. Mazario, K. Skinner and A.I. Basbaum, 2007. Anatomical and functional analysis of aquaporin 1, a water channel in primary afferent neurons. Pain, 131: 8-20.
- Ma, T., M. Hara, R. Sougrat, J.M. Verbavatz and A.S. Verkman, 2002. Impaired stratum corneum hydration in mice lacking epidermal water channel aquaporin-3. J. Biol. Chem., 277: 17147-17153.
- Tsukaguchi, H., C. Shayakul, U.V. Berger, B. Mackenzie and S. Devidas et al., 1998. Molecular characterization of a broad selectivity neutral solute channel. J. Biol. Chem., 273: 24737-24743.
- Saadoun, S., M.C. Papadopoulos, M. Hara-Chikuma and A.S. Verkman, 2005. Impairment of angiogenesis and cell migration by targeted aquaporin-1 gene disruption. Nature, 434: 786-792.
- Ozu, M., L. Galizia, C. Acuña and G. Amodeo, 2018. Aquaporins: More than functional monomers in a tetrameric arrangement. Cells, 7.
- Quiroga, G., G. Erice, L. Ding, F. Chaumont, R. Aroca and J.M. Ruiz‐Lozano, 2019. The arbuscular mycorrhizal symbiosis regulates aquaporins activity and improves root cell water permeability in maize plants subjected to water stress. Plant Cell Environ., 42: 2274-2290.
- D'Acierno, M., R.A. Fenton and E.J. Hoorn, 2025. The biology of water homeostasis. Nephrol. Dialysis Transplant., 40: 632-640.
- Afzal, Z., T. Howton, Y. Sun and M. Mukhtar, 2016. The roles of aquaporins in plant stress responses. J. Dev. Biol., 4.
- Verma, H., K. Devi, A.R. Baruah and R.N. Sarma, 2020. Relationship of root aquaporin genes, OsPIP1;3, OsPIP2;4, OsPIP2;5, OsTIP2;1 and OsNIP2;1 expression with drought tolerance in rice. Indian J. Genet. Plant Breed., 80: 50-57.
- Upadhyay, R., 2013. Plant Heavy Metal Stress. LAP Lambert Academic Publishing, Argentina, South America, ISBN: 9783659323447, Pages: 120.
- Upadhyay, R.K., 2014. Metal stress in plants: Its detoxification in natural environment. Braz. J. Bot., 37: 377-382.
- Kurowska, M.M., 2021. Aquaporins in cereals-important players in maintaining cell homeostasis under abiotic stress. Genes, 12.
- Knipfer, T. and W. Fricke, 2011. Water uptake by seminal and adventitious roots in relation to whole-plant water flow in barley (Hordeum vulgare L.). J. Exp. Bot., 62: 717-733.
- Sutka, M., G. Li, J. Boudet, Y. Boursiac, P. Doumas and C. Maurel, 2011. Natural variation of root hydraulics in arabidopsis grown in normal and salt-stressed conditions. Plant Physiol., 155: 1264-1276.
- Upadhyay, R., 2020. The Secret of Plants in the Environment. Notion Press, Chennai, India, ISBN: 9781648509209, Pages: 206.
- Ishibashi, K., S. Sasaki, K. Fushimi, S. Uchida and M. Kuwahara et al., 1994. Molecular cloning and expression of a member of the aquaporin family with permeability to glycerol and urea in addition to water expressed at the basolateral membrane of kidney collecting duct cells. Proc. Natl. Acad. Sci. U.S.A., 91: 6269-6273.
- Endo, M., R.K. Jain, B. Witwer and D. Brown, 1999. Water channel (aquaporin 1) expression and distribution in mammary carcinomas and glioblastomas. Microvasc. Res., 58: 89-98.
- Haj-Yasein, N.N., G.F. Vindedal, M. Eilert-Olsen, G.A. Gundersen and Ø. Skare et al., 2011. Glial-conditional deletion of aquaporin-4 (Aqp4) reduces blood-brain water uptake and confers barrier function on perivascular astrocyte endfeet. Proc. Natl. Acad. Sci. U.S.A., 108: 17815-17820.
- Pittock, S.J. and C.F. Lucchinetti, 2016. Neuromyelitis optica and the evolving spectrum of autoimmune aquaporin-4 channelopathies: A decade later. Ann. N. Y. Acad. Sci., 1366: 20-39.
- Bhattacharjee, H., B.P. Rosen and R. Mukhopadhyay, 2009. Aquaglyceroporins and Metalloid Transport: Implications in Human Diseases. In: Aquaporins, Beitz, E. (Ed.), Springer, Berlin Heidelberg, ISBN: 978-3-540-79885-9, pp: 309-325.
- Wang, J., L. Feng, Z. Zhu, M. Zheng, D. Wang, Z. Chen and H. Sun, 2015. Aquaporins as diagnostic and therapeutic targets in cancer: How far we are? J. Transl. Med., 13.
- Maugeri, R., G. Schiera, C.M. di Liegro, A. Fricano, D.G. Iacopino and I. di Liegro, 2016. Aquaporins and brain tumors. Int. J. Mol. Sci., 17.
- Pan, H., C.C. Sun, C.Y. Zhou and H.F. Huang, 2008. Expression of aquaporin-1 in normal, hyperplasic, and carcinomatous endometria. Int. J. Gynecology Obstet., 101: 239-244.
- Kaneko, K., K. Yagui, A. Tanaka, K. Yoshihara and K. Ishikawa et al., 2008. Aquaporin 1 is required for hypoxia-inducible angiogenesis in human retinal vascular endothelial cells. Microvasc. Res., 75: 297-301.
- Jiang, Y., 2009. Aquaporin-1 activity of plasma membrane affects HT20 colon cancer cell migration. IUBMB Life, 61: 1001-1009.
- Hara-Chikuma, M. and A.S. Verkman, 2005. Aquaporin‐3 functions as a glycerol transporter in mammalian skin. Biol. Cell, 97: 479-486.
- Zou, L.B., R.J. Zhang, Y.J. Tan, G.L. Ding and S. Shi et al., 2011. Identification of estrogen response element in the aquaporin-2 gene that mediates estrogen-induced cell migration and invasion in human endometrial carcinoma. J. Clin. Endocrinol. Metab., 96: E1399-E1408.
- Evans, J., A. Akhter, D. Carbone, M. Dikov and E. Tchekneva, 2017. P2.03b-082 AQP11 as a novel factor of lung cancer cell resistance to cisplatin. J. Thoracic Oncol., 12: S985-S986.
- Wang, G., F. Gao, W. Zhang, J. Chen, T. Wang, G. Zhang and L. Shen, 2012. Involvement of aquaporin 3 in Helicobacter pylori-related gastric diseases. PLoS ONE, 7.
- Dian, Z., Y. Sun, G. Zhang, Y. Xu and X. Fan et al., 2021. Rotavirus-related systemic diseases: Clinical manifestation, evidence and pathogenesis. Crit. Rev. Microbiol., 47: 580-595.
- Jennische, E., C.E. Eriksson, S. Lange, E. Trybala and T. Bergström, 2015. The anterior commissure is a pathway for contralateral spread of herpes simplex virus type 1 after olfactory tract infection. J. Neurovirol., 21: 129-147.
How to Cite this paper?
APA-7 Style
Upadhyay,
P., Upadhyay,
R. (2025). Aquaporins (AQPs): On Making Progress Towards their Role and Application in the Biological World. Asian Science Bulletin, 3(3), 254-264. https://doi.org/10.3923/asb.2025.254.264
ACS Style
Upadhyay,
P.; Upadhyay,
R. Aquaporins (AQPs): On Making Progress Towards their Role and Application in the Biological World. Asian Sci. Bul 2025, 3, 254-264. https://doi.org/10.3923/asb.2025.254.264
AMA Style
Upadhyay
P, Upadhyay
R. Aquaporins (AQPs): On Making Progress Towards their Role and Application in the Biological World. Asian Science Bulletin. 2025; 3(3): 254-264. https://doi.org/10.3923/asb.2025.254.264
Chicago/Turabian Style
Upadhyay, Pratiksha, and Rishikesh Upadhyay.
2025. "Aquaporins (AQPs): On Making Progress Towards their Role and Application in the Biological World" Asian Science Bulletin 3, no. 3: 254-264. https://doi.org/10.3923/asb.2025.254.264

This work is licensed under a Creative Commons Attribution 4.0 International License.